玉米ZmEXO70s基因家族鉴定及苗期耐热性关联分析
冯信 张春梅 王宏伟


摘要:【目的】鑒定玉米EXO70s(ZmEXO70s)基因家族成员,分析其遗传变异与玉米耐热性的关联性,为揭示其在玉米耐热方面的功能和分子机制打下基础。【方法】以拟南芥EXO70s(AtEXO70s)基因家族成员序列为参考,利用MaizeGDB和NCBI数据库从玉米全基因组中鉴定出ZmEXO70s基因家族成员,对其进行基因结构及系统进化分析,根据其在系统发育进化树上的位置对其进行系统命名,并基于转录组测序(RNA-Seq)数据分析ZmEXO70s基因家族成员在不同组织及非生物胁迫下的表达模式。最后,鉴定玉米自交群体AM368中ZmEXO70s基因SNP位点,结合玉米AM368群体苗期耐热存活率进行基因家族关联分析。【结果】从玉米全基因组序列中共鉴定出36个ZmEXO70s基因,分布在10条染色体上,长度为228~3726 bp,编码75~1241个氨基酸残基,蛋白分子量为8.6~136.6 kD,理论等电点(pI)介于4.5~9.9,大部分蛋白pI小于7.0。拟南芥、水稻和玉米的EXO70s蛋白被分为9组(A组~I组),其中ZmEXO70s蛋白在各组中均有分布。大多数ZmEXO70s基因不含内含子,只有少数基因含有内含子,且内含子数量不同。36个ZmEXO70s蛋白共含8种基序(Motif 1~Motif 8),主要集中在C端,说明这些蛋白端序列较保守,且Motif 1~Motif 8组成Exo70保守结构域。ZmEXO70s基因在不同组织中的表达水平均存在明显差异,其中ZmEXO70B3基因在雄穗、花药、叶片和花丝中高表达,ZmEXO70C1a基因只在雄穗和花药中高表达,ZmEXO70D2b基因在胚乳和萌发种子中低表达,ZmEXO70G1c基因在花丝和萌发种子中低表达。有23个ZmEXO70s基因至少可响应一种非生物胁迫,说明这些ZmEXO70s基因参与非生物胁迫响应过程。ZmEXO70D2a和ZmEXO70E1基因的遗传变异与玉米苗期耐热性具有显著相关性(P≤0.01)。【结论】ZmEXO70s基因家族成员在系统发育进化上较保守,其基因组复制事件可能发生在禾本科植物分化后,且大多数ZmEXO70s基因被保留,仅部分基因被丢失。ZmEXO70s基因可能在非生物胁迫响应过程中发挥重要作用,ZmEXO70D2a和ZmEXO70E1基因可作为调控玉米苗期耐热性重要候选基因。
关键词:
玉米;EXO70;基因家族鉴定;苗期耐热性;关联性分析;表达分析;胞泌复合体
中图分类号:
S513.035.2 文献标志码:
A 文章编号:2095-1191(2021)04-0867-12
Identification of ZmEXO70s gene family in maize and its correlation with heat tolerance at seedling stage
FENG Xin1, ZHANG Chun-mei1, WANG Hong-wei1,2
(1Agricultural College, Yangtze University, Jingzhou, Hubei 434025, China; 2Hubei Collaborative Innovation
Center for Grain Industry, Yangtze University, Jingzhou, Hubei 434025, China)
Abstract:【Objective】To identify the members of maize ZXO70s(ZmEXO70s) gene family and analyze the correlation between their genetic variation and maize heat tolerance, and lay the foundation for defining its functional and mole-cular mechanisms in maize heat tolerance. 【Method】With the sequences of Arabidopsis thaliana EXO70s(AtEXO70s) gene family members as reference, the members of the ZmEXO70s gene family were identified from the whole maize genome by MaizeGDB and NCBI databases, and the gene structure and phylogenetic analysis were carried out, and the members were systematically named according to their positions in the phylogenetic tree. Based on transcriptome sequencing (RNA-Seq) data, the expression patterns of ZmEXO70s gene family members under different tissues and abiotic stresses were analyzed. Finally, the SNP loci of ZmEXO70s gene in maize self-crossing population AM368 were identified, and gene family association analysis was conducted in combination with the heat resistance survival rate of maize AM368 at seedling stage. 【Result】A total of 36 ZmEXO70s genes were identified from the whole genome sequence of maize, distri-buted on 10 chromosomes, with lengths ranging from 228 to 3726 bp, encoding 75-1241 amino acids, molecular weight of proteins ranging from 8.6 to 136.6 kD, theoretical isoelectric point(pI) ranging from 4.5 to 9.9, and pI of most proteins was less than 7.0. The EXO70s proteins of A. thaliana, rice and maize were divided into 9 groups(groups A to I), with ZmEXO70s proteins distributed in each group. Most ZmEXO70s genes did not contain introns, and only a few did, with varying numbers of introns. The 36 ZmEXO70s proteins contained 8 motifs(Motif 1-Motif 8), which were mainly concentrated in the C terminal, indicating that the ZmEXO70s C terminal sequence was relatively conservative, and Motif 1-Motif 8 constituted the conservative domain of Exo70. The expression level of the ZmEXO70s gene varied greatly in different tissues. The ZmEXO70s gene was highly expressed in panicle and anther, leaf and filaments, ZmEXO70C1a gene was only highly expressed in panicle and anther, and ZmEXO70D2b gene was lowly expressed in endosperm and germinal seed. ZmEXO70G1c gene was lowly expressed in filaments and germinated seeds. Twenty-three ZmEXO70s genes could respond to at least one abiotic stress, suggesting that these ZmEXO70s genes were involved in the abiotic stress response process. The genetic variation of ZmEXO70D2a and ZmEXO70E1 was significantly correlated with maize seedling heat resistance(P≤0.01). 【Conclusion】The members of the ZmEXO70s gene family are relatively conserved in phylogenetic evolution, and the genome replication event may occur after the differentiation of Gramineae, and most of the ZmEXO70s genes are retained, while some of them are lost. ZmEXO70s may play an important role in abiotic stress response, ZmEXO70D2a and ZmEXO70E1 genes may be important candidate genes for the regulation of maize seedling heat tolerance.
Key words:
maize; EXO70; gene family identification; heat resistance at seedling stage; correlation analysis; expression analysis; secretion complex
Foundation item:
General Project of Hubei Natural Science Foundation(2019CFB616)
0 引言
【研究意义】胞吐是细胞中一类重要的囊泡运输,由高尔基体运输囊泡与细胞质膜融合,介导胞内物质分泌到细胞外及膜蛋白向质膜的转运过程(Zhang et al.,2010a;Heider and Munson,2012)。胞泌复合体负责将高尔基囊泡栓系到细胞质膜上(Sekere? et al.,2017;Kulich et al.,2018),是胞吐囊泡栓系过程中发挥关键作用的一类栓系因子,该复合体由8个蛋白亚基构成,其中EXO70是组成胞泌复合体功能的关键亚基,其决定胞泌复合体导向靶标膜的胞吐位点,参与细胞壁形成、细胞分泌和抗逆应答等重要细胞学过程(Sekere? et al.,2017;Kulich et al.,2018)。鉴定玉米(Zea mays L.)EXO70s基因(ZmEXO70s)家族成员,分析其进化及表达模式,尤其是探究其遗传变异与玉米耐热性的关系,对深入了解玉米响应高温胁迫的分子机制及发展玉米耐热性育种具有重要意义。【前人研究进展】目前关于植物胞泌复合体的研究主要集中在EXO70家族,其中在毛果杨(Populus trichocarpa)(Elias et al.,2003)、拟南芥(Arabidopsis thaliana)(Chong et al.,2010)、高粱(Sorghum bicolor)(Cvr?ková et al.,2012)、水稻(Oryza sativa)(Tu et al.,2015)、土豆(Symphytum tuberosum)(Du et al.,2018)和小麦(Triticum aestivum)(Zhao et al.,2018)等物种中已分别鉴定出31、23、22、41、29和75个EXO70蛋白,且部分EXO70蛋白的生物学功能已被验证。如拟南芥EXO70A1(AtEXO70H4)蛋白参与管状分子分化、组织(种皮、根毛和柱头乳突)发育及凯氏带形成(Kulich et al.,2010;Kalmbach et al.,2017;Vuka?inovi? and ?ársk?,2017),AtEXO70H4蛋白通过调节胼胝质和二氧化硅的分泌及积累以调控毛状体细胞壁的成熟(Chen et al.,2014),AtEXO70C1和AtEXO70C2蛋白可调控拟南芥花粉管成熟及其极性生长过程(Synek et al.,2017);水稻EXO70A1(OsEXO70A1)蛋白在维管束分化和矿物质营养同化过程中发挥重要作用(Tu et al.,2015)。此外,已有较多研究证实,EXO70s蛋白在植物与病原体的相互作用及对非生物胁迫的响应过程中发挥重要作用。如烟草(Nicotiana benthamiana)中2个EXO70B同源基因能沉默增加植株对晚疫病菌的感病性(Du et al.,2018);AtEXO70B1、AtEXO70B2和AtEXO70H1基因均参与植物免疫反应调控,其中AtEXO70B1基因缺失导致水杨酸积累,其突变体出现异位过敏反应并对多种病原体产生广谱抗性(Pe?enková et al.,2011;Kulich et al.,2013);AtEXO70B1和AtEXO70B2蛋白参与非生物应激反应,二者均为气孔运动的正向调节因子,如在干旱或甘露醇渗透胁迫下,通过依赖或不依赖脱落酸的信号传导来响应胁迫(Hong et al.,2016;Seo et al.,2016)。可见,EXO70s蛋白参与胞吐有关多种重要的生物学过程。【本研究切入点】虽然植物中存在大量的EXO70s基因,但目前仅对少数物种EXO70s基因进行生物学功能分析,鲜见有关ZmEXO70s基因家族的研究报道。因此,揭示ZmEXO70s基因家族成员数量、逆境胁迫响应机制及其与苗期耐热性的相关性,不仅有利于推进玉米耐热性的遗传改良,还有助于加深对该基因家族生物学功能的认识。【拟解决的关键问题】以AtEXO70s基因家族成员序列为参考,利用MaizeGDB和NCBI数据库从玉米全基因组中鉴定出ZmEXO70s基因家族成员,结合拟南芥和水稻的EXO70s蛋白进行系统发育进化分析及系统命名,并基于转录组测序(RNA-Seq)数据对ZmEXO70s基因家族成员在不同组织及非生物胁迫下的表达模式进行分析,探析ZmEXO70s基因的遗传变异与玉米耐热性的相关性,为揭示该基因家族在玉米生长发育和非生物胁迫响应中的调控机制提供理论依据,也为玉米耐热性分子育种提供基因资源。
1 材料与方法
1. 1 ZmEXO70s基因家族成员鉴定
从TAIR数据库中(https://www.arabidopsis.org/)查找23个AtEXO70s基因及其编码蛋白的相关信息;在RGAP7数据库中搜索OsEXO70s基因家族编码的蛋白序列。根据AtEXO70s和OsEXO70s蛋白序列,在Phytozome数据库中(https://phytozome.jgi.doe.gov/)對玉米B73基因组(AGPv3)进行BLASTp序列比对分析,并将获得的蛋白序列在NCBI的CDD(https://www.ncbi.nlm.nih.gov/cdd)进行蛋白结构预测,筛选出包含结构域pfam03081的序列,最终确定ZmEXO70s基因家族成员。
1. 2 ZmEXO70s基因结构及系统发育进化分析
使用ClustalX 2.0将ZmEXO70s基因编码区序列与其对应的参考基因序列进行比对,并使用CSDS 2.0进行基因结构分析。利用MEME和TBtools分析ZmEXO70s蛋白的保守基序(Motif),以Pfam在线分析ZmEXO70s蛋白的结构域,运用ClustalX 2.0比对ZmEXO70s、OsEXO70s和AtEXO70s蛋白序列,并以MEGA 7.0的邻接法(Neighbor-joining,NJ)构建系统发育进化树,参数是配对删除(Pairwise deletion)和1000次置换检验(Bootstrap)。
1. 3 ZmEXO70s基因在不同组织及非生物胁迫下的表达分析
利用玉米B73的RNA-Seq数据(Sekhon et al.,2011;Li et al.,2017)对ZmEXO70s基因在不同组织及非生物胁迫下的表达情况进行分析。以转录组分析通用的FPKM(Fragments per kilobase of transcript per million fragments mapped)值作为目的基因相对表达量。为方便统计,对每个表达数值取以2为底数的对数(log2)来绘制ZmEXO70s基因的表达热图。
1. 4 ZmEXO7基因遗传变异与玉米苗期耐热性的关联分析
ZmEXO70s耐热候选基因家族关联分析参照Li等(2013)的方法。该定位群体由全球代表性的368个玉米自交系群体(AM368)构成,含有55.6万高质量的SNP分子标记,其微等位基因频率(MAF)>0.05(Li et al.,2013)。使用TASSEL的2个统计模型(Bradbury et al.,2007),即一般线性模型(GLM)和混合线性模型(MLM)进行SNP分子标记与苗期耐热性的关联分析。其中,GLM是将群体结构考虑在内,结合基因型数据及表型数据进行关联分析;MLM则是将群体结构和亲缘关系均考虑在内,结合基因型数据及表型数据进行关联分析(Zhang et al.,2010b;Li et al.,2013)。
1. 5 AM368群体苗期耐热存活率表型
在长江大学农学院的步入式生长温室中对AM368群体自交系进行6次苗期耐热表型鉴定。以玉米苗期高温存活率为统计指标,试验设2次重复,采用随机区组设计。每个小钵子种植12棵玉米,称取相同重量的土壤基质,严格控制土壤基质的重量和水分,待幼苗生长至3叶1心期时,对其进行42 ℃高温胁迫处理。当玉米出现叶片萎蔫及植株死亡表型时温度降至25 ℃,并在25 ℃恢复7 d后统计各自交系的存活率。
2 结果与分析
2. 1 ZmEXO70s基因家族成员鉴定及蛋白理化性质分析结果
根据氨基酸序列相似性比对及NCBI数据库BLASTp序列比对等生物信息学分析,共鉴定出36个ZmEXO70s基因家族成员,其GenBank登录号、染色体位置、基因长度及编码蛋白的理化性质如表1所示。36个ZmEXO70s基因分布在10条染色体上,基因序列长度和氨基酸数目各不相同,其中ZmEXO70G1a基因序列长度为228~3726 bp,编码75~1241个氨基酸残基,蛋白分子量为8.6~136.6 kD,理论等电点(pI)为4.5~9.9,大部分蛋白理论等电点小于7.0。
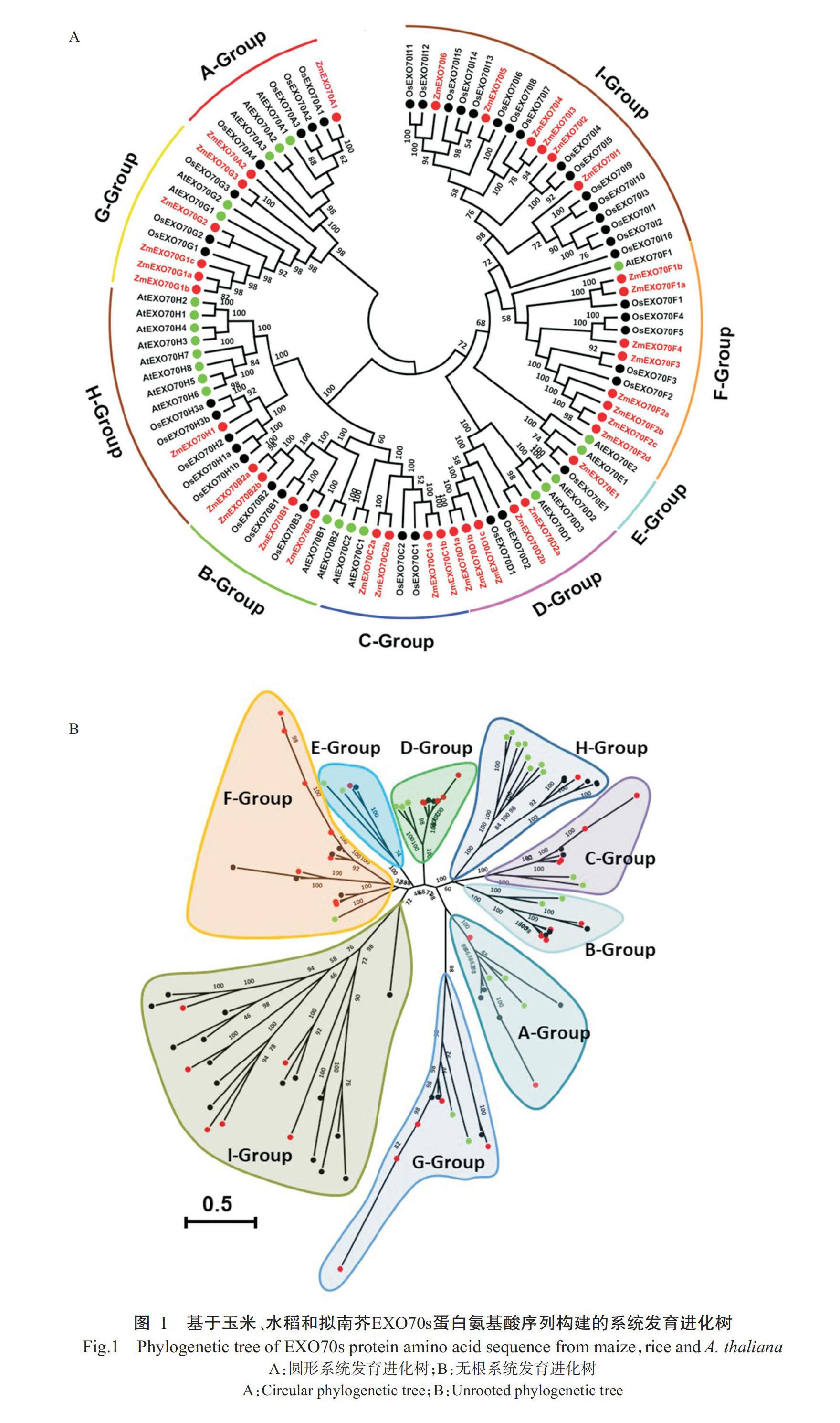
2. 2 ZmEXO70s蛋白系统发育进化分析结果
为了探究玉米与其他物种EXO70蛋白的同源进化关系,将已知的23个AtEXO70s蛋白(Chong et al.,2010)、41个OsEXO70s蛋白(Cvr?ková et al.,2012)和本研究鉴定出的36个ZmEXO70s蛋白进行系统发育进化分析,结果如图1-A所示。拟南芥、水稻和玉米的EXO70s蛋白可分为9组(A组~I组),各组数目不相同,以E组数目最少、I组数目最多。结合前人构建的AtEXO70s和OsEXO70s蛋白系统发育进化树(Chong et al.,2010;Cvr?ková et al.,2012),对ZmEXO70s基因家族成员进行系统命名。由图1-B可知,各组蛋白间有较高的同源性,十字花科植物拟南芥的AtEXO70s蛋白在不同组中均聚在同一小分支,OsEXO70s和ZmEXO70s蛋白则聚在其他小分支,究其原因可能是水稻和玉米同为禾本科植物,二者亲缘关系较近。由表2可知,ZmEXO70s蛋白在不同组中均有分布,数目较多的是F组和I组,分别含有8和6个;数目最少的是E组和H组,分别只有1个;I组中只包含ZmEXO70s和OsEXO70s蛋白,分别为6和16个;在亲缘关系较近的F组和I组中,ZmEXO70s与OsEXO70s蛋白数量相近,但明显多于AtEXO70s蛋白数量,其原因可能是十字花科植物进化为禾本科植物时水稻和玉米全基因组发生复制事件并保留下较多基因;H组中ZmEXO70s蛋白数量远少于AtEXO70s蛋白数量,其原因可能是H组中编码ZmEXO70s蛋白的基因在玉米全基因组复制事件中發生基因丢失现象(表2)。综上所述,ZmEXO70s蛋白在系统发育进化上较保守,玉米基因组复制事件可能发生在禾本科植物分化之后,且大部分编码ZmEXO70s蛋白的基因被保留,仅部分基因发生丢失。
2. 3 ZmEXO70基因结构与保守结构域分析结果
构建36个ZmEXO70s蛋白的系统发育进化树,如图2-A所示。36个ZmEXO70s分为9组,与表2相同。对ZmEXO70s基因结构进行序列结构分析,发现大多数基因不含有内含子,仅少数基因含内含子,且内含子数量不同,其中内含子数最多是ZmEXO70D1b基因,含15个内含子。为研究ZmEXO70s蛋白基序组成的多样性和保守性,利用MEME对其保守基序进行分析,结果如图2-B所示。36个ZmEXO70s蛋白共含8种基序(Motif 1~Motif 8),且各基序主要集中在基因的C端,暗示ZmEXO70s蛋白C端序列具有较高的保守性。从基序种类来看,B组、D组和E组的ZmEXO70s蛋白保守性更高,包含全部基序,其他组ZmEXO70s在基序种类分布上存在明显差异,部分成员出现不同种类基序丢失现象,如ZmEXO70G1a蛋白只含有Motif 1,ZmEXO70F2c蛋白只含有Motif 3。此外,同一组内的ZmEXO70s蛋白含有不同基序,如ZmEXO70A2蛋白缺少Motif 1、Motif 2和Motif 3,但ZmEXO70A1蛋白只含有Motif 1、Motif 2和Motif 3;ZmEXO70F2b和ZmEXO70F2d蛋白只含有Motif 2和Motif 3,ZmEXO70F2c蛋白只含有Motif 3。虽然8种基序的功能尚不明确,但通过Pfam分析保守结构域,结果显示Motif 1~Motif 8组成Exo70保守结构域(图2-C)。
已有研究证实EXO70蛋白C端含有多个高度保守且带有正电荷的碱性氨基酸,与带有负电荷的磷脂酰肌醇4,5-二磷酸發生互作,促进胞泌复合体其他亚基募集至胞吐部位(Pe?enková et al.,2020)。为此,本研究将ZmEXO70s蛋白C端可能与质膜脂质互作有关的氨基酸序列进行多重比对分析,结果如图3所示。与质膜脂质互作有关的氨基酸序列由65~75个氨基酸组成,包含11个碱性氨基酸(蓝色星号)和9个酸性氨基酸(红色星号),其中有多个氨基酸位点相对保守。除ZmEXO70A1、ZmEXO70G1a、ZmEXO70F2b、ZmEXO70F2c和ZmEXO70F2d不含该段氨基酸序列外,其余31个ZmEXO70s蛋白C端均含该段氨基酸序列。在11个碱性氨基酸中,除第1和10个碱性氨基酸以外,其余9个碱性氨基酸在ZmEXO70s蛋白中保守性较高。在9个酸性氨基酸中,除第1、5和6个以外,其余6个酸性氨基酸在ZmEXO70s中也具有较高的保守性。但目前对这些保守的酸性和碱性氨基酸在Exo70结构域中的功能尚不清楚,推测部分位点的氨基酸差异影响ZmEXO70s蛋白脂质结合能力,进而影响蛋白在细胞中的定位,或是ZmEXO70s蛋白功能差异的重要决定因素。
2. 4 ZmEXO70s基因在不同组织及非生物胁迫下的表达分析结果
利用玉米B73的RNA-Seq数据(Sekhon et al.,2011),对36个ZmEXO70s基因在15个组织中的表达特性进行分析,结果如图4-A所示。组间和组内的ZmEXO70s基因表达水平均存在明显差异,其中ZmEXO70A2、ZmEXO70F1a和ZmEXO70AF1b基因表达水平较高,其次是ZmEXO70B1、ZmEXO70B2a、ZmEXO70B2b、ZmEXO70D1a、ZmEXO70D1c、Zm-EXO70D2a、ZmEXO70D2b、ZmEXO70E1、ZmEXO70-F1a、ZmEXO70F1b、ZmEXO70F2a、ZmEXO70F2b、ZmEXO70G1b、ZmEXO70G1c和ZmEXO70G2等15个ZmEXO70s基因,表达水平也较高,推测这些Zm-EXO70s基因是玉米生长发育中的重要调控基因;剩余基因的表达水平相对较低。值得注意的是,Zm-EXO70B3基因在雄穗、花药、叶片和花丝中高表达,ZmEXO70C1a基因仅在雄穗和花药中高表达,ZmEXO70D2b基因在胚乳和萌发种子中低表达,ZmEXO70G1c基因在花丝和萌发种子中低表达。
利用Li等(2017)的RNA-Seq数据(PRJNA244661和PRJNA335771)分析ZmEXO70s基因在干旱、高温、高盐和低温等非生物胁迫下的表达模式,结果如图4-B所示。在36个ZmEXO70s基因中,有23个ZmEXO70s基因至少可响应一种非生物胁迫,说明这些ZmEXO70s基因响应非生物胁迫,其余13个ZmEXO70s基因可能表达量极低,在RNA-Seq数据中未检测出。干旱胁迫下,ZmEXO70D1和ZmEXO70I2基因上调表达,ZmEXO70A1、ZmEXO70B3、ZmEXO-70D1a、ZmEXO70G1c和ZmEXO70I6基因下调表达。高温胁迫下,ZmEXO70s基因表现出不同的表达模式,如ZmEXO70D2b、ZmEXO70E1、ZmEXO70F2a和ZmEXO70F4基因上调表达,而ZmEXO70F1b、ZmEXO70H1和ZmEXO70I6基因下调表达。高盐胁迫和低温胁迫下,大多数ZmEXO70s基因被诱导上调表达,仅少数基因表达受到抑制,如ZmEXO70G1c和ZmEXO70H1基因在高盐和低温胁迫下均下调表达,ZmEXO70A1和ZmEXO70A1基因在低温胁迫下的表达轻微下调。推测大多数ZmEXO70s基因在非生物胁迫响应过程中发挥重要调控作用。
2. 5 ZmEXO70s基因遗传变异与玉米苗期耐热性的关联分析结果
利用基因家族关联分析方法检测ZmEXO70s基因的遗传变异与玉米苗期耐热性是否关联。玉米苗期耐热性是以玉米3叶1心期植株经持续高温胁迫下植株存活率为衡量指标。首先对AM368群体ZmEXO70s基因进行SNP分析,结果如表3所示。有31个ZmEXO70s基因含SNP位点,平均SNP位点数为21个,其中以ZmEXO70F2a基因SNP位点数最多,达85个。随后采用2种统计模型来分析基因型与表型间的相关性,结果如图5所示。通过GLM分析发现,ZmEXO70D2a、ZmEXO70E1和ZmEXO70I2基因的遗传变异与苗期耐热性存在显著相关性(P≤0.01)。通过MLM分析发现,ZmEXO70D2a和ZmEXO70E1基因分别有2个SNP位点与苗期耐热性呈极显著相关(P≤0.001,下同)。进一步分析发现,ZmEXO70E1基因的2个SNP位点位于其5"-非编码区(UTR)(图6-A);ZmEXO70D2a基因的2个SNP位点位于其编码区(图6-B)。
3 讨论
玉米是目前我国种植面积最大的作物。随着测序技术不断发展和玉米全基因组测序的完成,为玉米基因家族鉴定、功能基因挖掘及基因功能研究等提供极有利的先决条件。虽然现已证实不同植物中EXO70s蛋白参与细胞壁形成、细胞分泌和抗逆应答等重要细胞学过程(Sekere? et al.,2017;Kulich et al.,2018),但目前鲜见有关ZmEXO70s基因家族的研究报道。本研从玉米基因组中共鉴定出36个ZmEXO70s基因,说明EXO70s蛋白在玉米中与其他植物一样,均为多拷贝。植物EXO70s蛋白多拷贝现象可能是因为植物具有多种多样的细胞壁及复杂的液泡结构,需要多种胞泌复合体满足其适应环境的需要,进而演化出不同基因特有的调控机制(Elias et al.,2003;Chong et al.,2010)。ZmEXO70s基因家族成员数(36个)与高梁Ex070s(SbEXO70s)基因家庭数目(31个)相当,但比AtEXO70s基因家族成员数(23个)多,比OsEXO70s基因家族成员数(41个)少,可能是由于禾本科植物(水稻、玉米和高粱)在物种形成和进化过程中发生了全基因组复制和二倍体化,从而导致其EXO70s基因家族成员数目比十字花科的拟南芥多,且玉米和高粱在进化过程中经历染色体的丢失和融合,导致二者的EXO70s基因家族成员数目均比水稻少。
基因表达模式暗示其所行使的生物学功能。已有研究表明,胞泌复合体负责将高尔基囊泡栓系到细胞质膜上,其基因缺失或突變均会导致其失去该生物学功能(Koumandou et al.,2007;Drdová et al.,2013)。本研究对36个ZmEXO70s基因在15个组织中的表达水平进行分析,结果显示大多数Zm-EXO70s基因在玉米组织中均有表达,推测这些ZmEXO70s基因是玉米生长发育中重要调控基因,其中,ZmEXO70B3和ZmEXO70C1a基因在雄穗和花药中高表达。Li等(2010)也研究发现,AtEXO70C1基因在成熟花粉中高效表达,当其发生突变后会导致花粉管生长迟缓,花粉的传递效率下降。可见,植物EXO70s基因家族中部分成员可能参与调控植株花粉发育过程。此外,已有研究表明,囊泡运输相关蛋白通过参与非生物胁迫下的质膜修复增加植物对低温、高盐等非生物胁迫的耐受性(Yamazaki et al.,2008;Kim and Bassham,2011;Drakakaki et al.,2012),但针对胞泌复合体在非生物胁迫应答响应过程的功能研究较少。本研究通过分析ZmEXO70s基因在干旱、高温、高盐和低温等非生物胁迫下的表达模式,结果显示36个ZmEXO70s基因中有23个ZmEXO70s至少响应一种非生物胁迫,暗示ZmEXO70s基因可能参与调控植物非生物胁迫响应过程,为进一步阐明植物胞泌复合体功能的分子机制提供理论依据。
虽然玉米来源于热带,但仍对高温环境敏感,当气温超过35 ℃时,玉米植株整个生命周期(营养生长和生殖生长)均会受到影响(牛丽等,2015;Shi et al.,2017;石江等,2018),如春播玉米在抽雄吐丝期、夏播青储玉米在幼苗出土期遭遇高温热害的问题在我国玉米生产上表现越来越突出(Hansen et al.,2001;Lesk et al.,2016)。虽然目前没有研究直接证实,囊泡运输与植物耐热性相关,但已有研究证实囊泡运输参与气孔运动过程,如负调控拟南芥光诱导的气孔开放因子ROP2和RIC7,通过抑制EXO70B1蛋白来阻止气孔过度开放,说明EXO70B1通过介导囊泡运输间接调控气孔开放(Hong et al.,2016)。高温胁迫下植物通过增大叶片气孔开度,加快水分散失从而降低叶片温度以适应高温环境。由此可见,植物可能通过囊泡运输调控气孔运动从而响应高温胁迫。本研究通过基因家族关联分析发现,ZmEXO70D2a和ZmEXO70E1基因的遗传变异与玉米苗期耐热性呈极显著相关,且RNA-Seq数据分析结果显示,ZmEXO70D2a和ZmEXO70E1基因的表达受高温胁迫分别被抑制或诱导,故推测ZmEXO70D2a和ZmEXO70E1基因参与玉米高温胁迫响应过程,可作为提高玉米耐热性的重要候选基因。但高温胁迫下ZmEXO70D2a和ZmEXO70E1是否调控玉米叶片气孔的动态仍有待进一步研究。
4 结论
ZmEXO70s基因家族成员在系统发育进化上较保守,其基因组复制事件可能发生在禾本科植物分化后,且大多数ZmEXO70s基因被保留,仅部分基因被丢失。ZmEXO70s基因可能在非生物胁迫响应过程中发挥重要作用,ZmEXO70D2a和ZmEXO70E1基因可作为调控玉米苗期耐热性的重要候选基因。
参考文献:
牛丽,刘源,于康珂,刘荣花,李潮海,刘天学. 2015. 玉米杂交种苗期耐热性评价[J]. 玉米科学,23(1):107-114. doi:10. 13597/j.cnki.maize.science.2015.01.018. [Niu L,Liu Y,Yu K K,Liu R H,Li C H,Liu T X. 2015. Evaluation of heat-tolerance of maize hybrids at seedling stage[J]. Journal of maize sciences,23(1):107-114.]
石江,赵琳,朱月清,楼旭平,余建忠,阮松林,陈文岳. 2018. 玉米幼苗叶片响应热胁迫的蛋白质组学分析[J]. 浙江农业学报,30(6):893-908. doi:10.3969/j.issn.1004-1524. 2018.06.03. [Shi J,Zhao L,Zhu Y Q,Lou X P,Yu J Z,Ruan S L,Chen W Y. 2018. Proteomic analysis of maize seedling leaves in response to heat stress[J]. Acta Agriculturae Zhejiangensis,30(6):893-908.]
Bradbury P J,Zhang Z W,Kroon D E,Casstevens T M,Ramdoss Y,Buckler E S. 2007. TASSEL:Software for associa-tion mapping of complex traits in diverse samples[J]. Bioinformatics,23(19):2633-2635. doi:10.1093/bioinforma-tics/btm308.
Chen C H,Liu M,Jiang L,Liu X F,Zhao J Y,Yan S S,Yang S,Ren H Z,Liu R Y,Zhang X L. 2014. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.)[J]. Journal of Experimental Botany,65(17):4943-4958. doi:10.1093/jxb/eru258.
Chong Y T,Gidda S K,Sanford C,Parkinson J,Mullen R T,Goring D R. 2010. Characterization of the Arabidopsis thaliana exocyst complex gene families by phylogenetic,expression profiling,and subcellular localization studies[J]. New Phytologist,185(2):401-419. doi:10.2307/256 09624.
Cvr?ková F,Grunt M,Bezvoda R,Hála M,Kulich I,Rawat A,Zársk? V. 2012. Evolution of the land plant exocyst complexes[J]. Frontier in Plant Science,3:159. doi:10.3389/ fpls.2012.00159.
Drakakaki G,van de Ven W,Pan S Q,Miao Y S,Wang J Q,Keinath N F,Weatherly B,Jiang L W,Schumacher K,Hicks G,Raikhel N. 2012. Isolation and proteomic analysis of the SYP61 compartment reveal its role in exocytic trafficking in Arabidopsis[J]. Cell Research,22(2):413-424. doi:10.1038/cr.2011.129.
Drdová E J,Synek L,Pe?enková T,Hála M,Kulich I,Fowler J E,Murphy A S,Zársk? V. 2013. The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis[J]. The Plant Journal,73(5):709-719. doi:10.1111/tpj.12074.
Du Y,Overdijk E J R,Berg J A,Govers F,Bouwmeester K. 2018. Solanaceous exocyst subunits are involved in immunity to diverse plant pathogens[J]. Journal of Experimental Botany,69(3):655-666. doi:10.1093/jxb/erx442.
Elias M,Drdova E,Ziak D,Bavlnka B,Hala M,Cvrckova F,Soukupova H,Zarsky V. 2003. The exocyst complex in plants[J]. Cell Biology International,27(3):199-201. doi:
10.1016/s1065-6995(02)00349-9.
Hansen P J,Drost M,Rivera R M,Paula-Lopes F F,Al-Katanani Y M,Krininger C E,Chase C C J. 2001. Adverse impact of heat stress on embryo production:Causes and strategies for mitigation[J]. Theriogenology,55(1):91-103. doi:10.1016/s0093-691x(00)00448-9.
Heider M R,Munson M. 2012. Exorcising the exocyst complex[J]. Traffic,13(7):898-907. doi:10.1111/j.1600-0854. 2012.01353.x.
Hong D,Jeon B W,Kim S Y,Hwang J U,Lee Y. 2016. The ROP2-RIC7 pathway negatively regulates light-induced stomatal opening by inhibiting exocyst subunit EXO70B1 in Arabidopsis[J]. New Phytologist,209(2):624-635. doi:10.1111/nph.13625
Kalmbach L,Hématy K,De Bellis D,Barberon M,Fujita S,Ursache R,Daraspe J,Geldner N. 2017. Transient cell-specific EXO70A1 activity in the CASP domain and casparian strip localization[J]. Nature Plants,3:17058. doi:10.1038/nplants.2017.58.
Kim S J,Bassham D C. 2011. TNO1 is involved in salt tolera-nce and vacuolar trafficking in Arabidopsis[J]. Plant Physiology,156(2):514-526. doi:10.1104/pp.110.168963.
Koumandou V L,Dacks J B,Coulson R M,Field M C. 2007. Control systems for membrane fusion in the ancestral eukaryote;evolution of tethering complexes and SM proteins[J]. BMC Evolutionary Biology,7:29. doi:10.1186/1471-2148-7-29.
Kulich I,Cole R,Drdová E,Cvrcková F,Soukup A,Fowler J,Zársk? V. 2010. Arabidopsis exocyst subunits SEC8 and EXO70A1 and exocyst interactor ROH1 are involved in the localized deposition of seed coat pectin[J]. New Phytologist,188(2):615-625. doi:10.1111/j.1469-8137.2010. 03372.x.
Kulich I,Pe?enková T,Sekere? J,Smetana O,Fendrych M,Foissner I,H?ftberger M,Zársk? V. 2013. Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole[J]. Traffic,14(11):1155-1165. doi:10.1111/tra.12101.
Kulich I,Vojtíková Z,Sabol P,Ortmannová J,Neděla V,Tihla?íková E,?ársk? V. 2018. Exocyst subunit EXO70H4 has a specific role in callose synthase secretion and silica accumulation[J]. Plant Physiology,176(3):2040-2051. doi:10.1104/pp.17.01693.
Lesk C,Rowhani P,Ramankutty N. 2016. Influence of extreme weather disasters on global crop production[J]. Nature,529(7584):84-87. doi:10.1038/nature16467.
Li H,Peng Z Y,Yang X H,Wang W D,Fu J J,Wang J H,Han Y J,Chai Y C,Guo T T,Yang N,Liu J,Warburton M L,Cheng Y B,Hao X M,Zhang P,Zhao J Y,Liu Y J,Wang G Y,Li J S,Yan J B. 2013. Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels[J]. Nature Genetics,45(1):43-50. doi:10.1038/ng.2484.
Li P C,Cao W,Fang H M,Xu S H,Yin S Y,Zhang Y Y,Lin D Z,Wang J N,Chen Y F,Xu C W,Yang Z F. 2017. Transcriptomic profiling of the maize(Zea mays L.) leaf response to abiotic stresses at the seedling stage[J]. Frontiers in Plant Science,8:290. doi:10.3389/fpls.2017.00290.
Li S P,van Os G M,Ren S C,Yu D L,Ketelaar T,Emons A M,Liu C M. 2010. Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis[J]. Plant Physiology,154(4):1819-1830. doi:10.1104/pp.110.164178.
Pe?enková T,Hála M,Kulich I,Kocourková D,Drdová E,Fendrych M,Toupalová H,Zársky V. 2011. The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction[J]. Journal of Experimental Botany,62(6):2107-2116. doi:10.1093/jxb/erq402.
Pe?enková T,Potocká A,Potock? M,Ortmannová J,Drs M,Janková Drdová E,Pejchar P,Synek L,Soukupová H,?ársk? V,Cvr?ková F. 2020. Redundant and diversified roles among selected Arabidopsis thaliana EXO70 paralogs during biotic stress responses[J]. Frontiers in Plant Science,11:960. doi:10.3389/fpls.2020.00960.
Sekere? J,Pejchar P,?antr??ek J,Vuka?inovi? N,?ársk? V,Potock? M. 2017. Analysis of exocyst subunit EXO70 family reveals distinct membrane polar somains in tobacco pollen tubes[J]. Plant Physiology,173(3):1659-1675. doi:10.1104/pp.16.01709.
Sekhon R S,Lin H N,Childs K L,Hansey C N,Buell C R,de Leon N,Kaeppler S M. 2011. Genome-wide atlas of transcription during maize development[J]. The Plant Journal,66(4):553-563. doi:10.1111/j.1365-313X.2011.045 27.x.
Seo D H,Ahn M Y,Park K Y,Kim E Y,Kim W T. 2016. The N-terminal UND motif of the Arabidopsis U-Box E3 ligase PUB18 is critical for the negative regulation of ABA-mediated stomatal movement and determines its ubiquitination specificity for exocyst subunit Exo70B1[J]. The Plant Cell,28(12):2952-2973. doi:10.1105/tpc. 16.00347.
Shi J,Yan B Y,Lou X P,Ma H S,Ruan S L. 2017. Comparative transcriptome analysis reveals the transcriptional alterations in heat-resistant and heat-sensitive sweet maize(Zea mays L.) varieties under heat stress[J]. BMC Plant Biology,17(1):26. doi:10.1186/s12870-017-0973-y.
Synek L,Vuka?inovi? N,Kulich I,Hála M,Aldorfová K,Fendrych M,?ársk? V. 2017. EXO70C2 is a key regulatory factor for optimal tip growth of pollen[J]. Plant Physio-logy,174(1):223-240. doi:10.1104/pp.16.01282.
Tu B,Hu L,Chen W L,Li T,Hu B H,Zheng L,Lü Z,You S J,Wang Y P,Ma B T,Chen X W,Qin P,Li S G. 2015. Disruption of OsEXO70A1 causes irregular vascular bundles and perturbs mineral nutrient assimilation in rice[J]. Scientific Reports,5:18609. doi:10.1038/srep18609.
Vuka?inovi? N,?ársk? V. 2016. Tethering complexes in the Arabidopsis endomembrane system[J]. Frontiers in Cell and Development Biology,4:46. doi:10.3389/fcell.2016. 00046.
Yamazaki T,Kawamura Y,Minami A,Uemura M. 2008. Calcium-dependent freezing tolerance in Arabidopsis involves membrane resealing via synaptotagmin SYT1[J]. The Plant Cell,20(12):3389-3404. doi:10.1105/tpc.108.062679.
Zhang Y,Liu C M,Emons A M,Ketelaar T. 2010a. The Plant exocyst[J]. Journal of Integrative Plant Biology,52(2):138-146. doi:10.1111/j.1744-7909.2010.00939.x.
Zhang Z W,Ersoz E,Lai C Q,Todhunter R J,Tiwari H K,Gore M A,Bradbury P J,Yu J M,Arnett D K,Ordovas J M,Buckler E S. 2010b. Mixed linear model approach adapted for genome-wide association studies[J]. Nature Genetics,42(4):355-360 doi:10.1038/ng.546.
Zhao J,Zhang X,Wan W T,Zhang H,Liu J,Li M L,Wang H Y,Xiao J,Wang X. 2018. Identification and characterization of the EXO70 gene family in polyploid wheat and related species[J]. International Journal of Molecular Scien-ces,20(1):60. doi:10.3390/ijms20010060.
(責任编辑 陈 燕)
猜你喜欢 玉米 我的玉米送给你小天使·二年级语数英综合(2019年10期)2019-11-08为什么玉米长“胡须”好孩子画报(2019年6期)2019-06-30凶器是玉米小猕猴智力画刊(2019年3期)2019-04-19玉米的胡须数学大王·低年级(2018年3期)2018-03-27碾玉米作文大王·低年级(2017年10期)2017-10-28中国玉米供需平衡表农产品市场周刊(2017年4期)2017-03-03最棒的玉米作文周刊·小学二年级版(2016年23期)2016-09-14最棒的玉米当代学生·时政版(2009年9期)2009-10-123月16日各地玉米报价等农家顾问(2009年4期)2009-04-29