植物赤霉素受体GID1基因的分析
唐文芳 徐升胜 龙雯虹

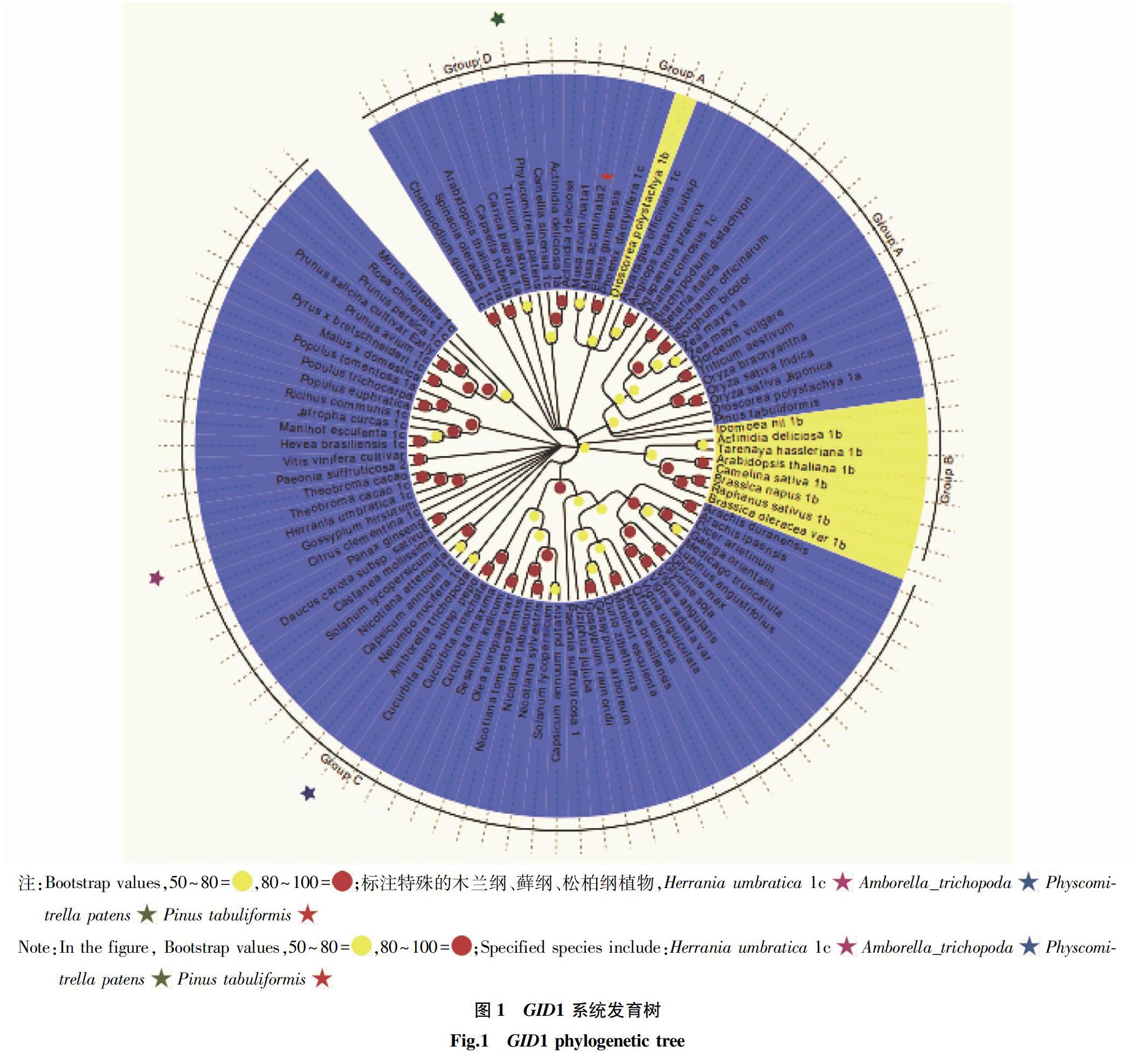
摘要 从NCBI数据库在线网站搜集了赤霉素受体基因核苷酸序列,分析了所获基因的进化关系。结果表明,植物的98个GID1基因拥有共同的根系,进化为Group A、Group B、 Group C和 Group D 4组。Group A GID1基因在单子叶植物纲中没有GID1a、GID1b、GID1c分类现象,在双子叶植物纲中基因有简单的Group B GID1b,Group C GID1a,且GID1c聚类现象;Group D由被子植物门单子叶植物和双子叶植物及苔藓植物门小立苔藓植物的基因组成;GID1基因组在进化过程中可能出现复制突变,所有GID1基因具有相同的保守区域,可利用基因之间的保守性来克隆未知基因;102条序列归属于91种植物,尚有更多单子叶植物、裸子植物和蕨类植物的GID1基因有待克隆。
关键词 赤霉素;GID1基因;系统发育重建
中图分类号 Q-943.2 文献标识码 A 文章编号 0517-6611(2021)12-0098-05
doi:10.3969/j.issn.0517-6611.2021.12.025
开放科学(资源服务)标识码(OSID):
Analysis of Gibberellin Receptor GID1 Gene in Plants
TANG Wen fang, XU Sheng sheng, LONG Wen hong
(Yunnan Agricultural University, Kunming, Yunnan 650201)
Abstract The nucleotide sequence of gibberellin receptor gene was collected from the NCBI database online website, and the evolutionary relationship of the obtained gene was analyzed. The results showed that 98 GID1 genes in plants had the same root system, which evolved into four groups:
Group A, Group B, Group C and Group D. Group A GID1 gene has no classification phenomenon of GID1a, GID1b and GID1c in monocotyledons. In dicotyledons, there are simple Group B GID1b, Group C GID1a and GID1c clustering phenomenon. Group D is composed of GID1 genes of Monocotyledoneae and Dicotyledoneae of the Angiospermae and Bryophyta Bryopsida of the plants. This suggests that the GID1 genome is replicating during the evolutionary process. All GID1 genes share the same conserved region, which can be used to clone unknown genes. Phylogenetic analysis of 102 GID1 genes of 91 plant species shows that more GID1 genes of monocotyledons, gymnosperms and ferns need to be cloned.
Key words Gibberellin;GID 1 genes;Phylogenetic reconstruction
赤霉素(gibberellins,GAs)是重要的植物激素之一,它參与植物生长发育的过程,包括种子萌发、茎叶的发育、花及果实的发育等[1-2]。有研究表明,赤霉素促进植物生长发育的作用较其他四大激素大,促进作用不会发生超最适浓度的抑制作用[3]。赤霉素通过生物合成和信号转导途径来影响植物生长发育,增加细胞的伸长,提高生物量,促进果实发育。信号转导途径,使植物产生赤霉素效应[4]。该过程必须通过赤霉素信号通路来实现,目前已发现130余种赤霉素类物质,但仅有少数几种具有生物活性,分别为GA1、GA3、GA4和GA7,主要用来调控植物生长发育的是GA3和GA7的混合物,它们的调控作用通过与Gibberellin Insensitive Dwarf 1(GID1)结合的亲和度来反映[5]。赤霉素受体GID1不论在蛋白水解途径还是非蛋白水解途径中都有特殊功能[4],每个受体基因在不同蛋白水解途径中对植物种子发芽、根茎叶的生长及繁殖起着不同的作用[3,6]。
GID1基因受体有3种类型,遗传分析表明,GA在信号转导中具有3种特异性功能[6],3种GA受体GID1A、GID1B和GID1C通过GA/GID1刺激破坏GA反应的Dell抑制因子而积极调节GA反应[7],比如GID1基因在种子萌发过程中GID1BmRNA和DELLA蛋白有较高的亲和力,在泛素化降解蛋白中不影响 DELLA的活性,表达水平较高[6];GID1A和GID1C保留典型的GA信号作用,GID1B的表达偏向于地下组织和干豆类种子[8]。以野生型拟南芥种子萌发为例,GID1B可以作为上游负调控因子GID1C,休眠种子萌发的正调控因子,暗萌发的负调节因子;GID1B和GID1A在一定程度上可以作为GID1C的上游负调控因子,由于GID1 3突变体在黑暗中不能萌发,因此GID1C可能是暗萌发的关键下游正调控因子[9]。3种受体GID1在功能上相互分离又相互补充,并且每个基因可能在不同的组织器官和不同发育时期发挥特殊功能[9]。
目前,对GID1基因的研究广泛,部分物种的基因序列已被克隆出,但对植物的GID1基因进化关系仍缺乏较为系统的研究。为明确不同物种GID1在发育中的关系,笔者搜集了已登录的GID1基因序列,分析GID1基因的进化关系,旨在为深入研究GID1基因的克隆现状提供参考。
1 材料与方法
1.1 GID1序列的筛选
NCBI(https://www.ncbi.nlm.nih.gov/gene/)数据库中输入关键词“gid1”,搜索出相关序列,找出其中GID1基因序列,Blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)基因序列在线比对筛选,得到拟进行系统进化分析的赤霉素受体基因序列。在《中国植物志》官网查对这些基因拉丁名对应的中文所属,在线网址:(http://www.iplant.cn/frps)。
1.2 重建GID1系统发育树
首先把筛选好的基因序列用Editseq MegAlign Clustalx(1.83)软件整合成FASTA格式,然后把FASTA文件导入Mega7.0点击新窗口“TA”Mega7.0 Sequence Data Explorer进行多基因序列分析,Mega7.0 Sequence Data Explorer窗口点击“TA”按钮后,再点击“C”按钮,通过比较序列间的保守程度,淘汰一些不合群的序列后最小化窗口,回到Mega7.0 Phylogeny窗口使用邻位相连法(Neighbor joining,NJ)导入文件构建系统进化树,NJ 法中的稳健性采用自展内部分枝法(Boot strapping)进行评定,设置Bootstrap Replication值,密码子位置选用1+2+3+非编码,删除每个序列对应的所有不明确的位置,最后得到完整的系统发育树。Evolview在线编辑系统发育树,编辑网址:https://evolgenius.info//evolview v2/#mytrees/DEMOS/yeast%20duplications。
1.3 GID1保守序列分析
在Mega7.0 Sequence Data Explorer窗口对基因序列进行保守性分析。
2 结果与分析
2.1 植物GID1基因登录情况
NCBI Gene数据库中输入关键词“gid1”,共搜索出2 436个基因,其中关于GID1的有114个,进行Blast基因比对,最终获得102个基因序列(表1)。102个基因的拉丁文登录名查对《中国植物志》得知,该研究搜集的基因序列分属于91种植物,分别为68个双子叶植物,20个单子叶植物,1个松柏纲植物,1个藓纲植物,1个木兰纲植物。分析植物种属分布情况可以得出GID1基因的登录情况,被子植物门占基因总登录物种的97.80%,双子叶植物纲基因占基因总登录的74.73%,占被子植物门的77.53%;单子叶植物纲基因占总登录基因的21.97%,占被子植物门的22.47%;裸子植物门占基因总登录的1.10%;蕨类植物占总登录的1.10%;苔藓植物占总登录的1.10%。这说明GID1基因的研究集中于被子植物门,其他植物GID1基因的登录情况较少。
2.2 GID1的系统进化
2.2.1 GID1 系统发育树。
基因序列整合导入Mega7.0后,分析删除不必要基因序列(序列11、序列17、序列18和序列76)后,邻近法Bootstrap Replication值设置为1 000构建完整的进化树(图1)。图1显示,GID1拥有共同的根系、共同的祖先,进化为顺时针方向。根据植物学分类及GID1s命名方式将98个GID1基因順时针方向分为4个组,分别为Group A、 GroupB、 GroupC、 GroupD。Group A GID1基因序列分属于被子植物门单子叶植物和裸子植物门松纲植物;Group B分布被子植物门双子叶植物GID1b序列;Group C GID1序列主要分布被子植物门双子叶植物和双子叶木兰纲植物的基因,分化出两大分枝(如图2蓝色枝),上枝基因序列间自展值在80%以上,且80%以上为GID1c,下枝GID1基因序列的自展值达到80%的仅占50%。Group B双子叶植物GID1b与Group A中单子叶植物GID1自展值超过50%。Group D是被子植物门单子叶植物和双子叶植物及苔藓植物门苔藓植物的基因组成。
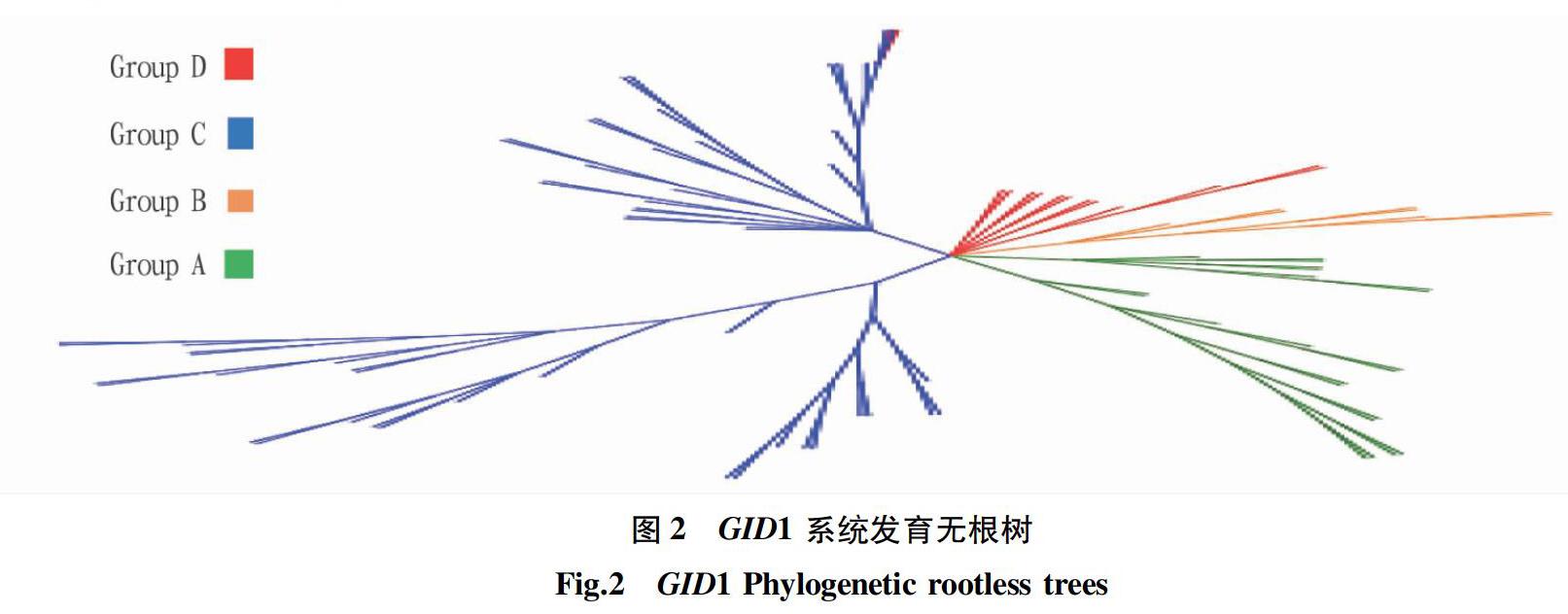
从GID1系统发育无根树(图2)可以看出,GID1树枝分散,分散成主要四大组枝,各大组枝分化明显,表明这91种植物之间的基因存在差异。
2.2.2 基因保守序列。
从图3可以看出,GID1基因序列的保守序列为GGT TCC CTG AAC CTC ATC T TTA C TCT TCG TCC GGA TGG CAC ATT CAA CG AA AGC。
通过标准方式分析基因的核酸,比较GID1基因序列的保守碱基数来分析各组基因之间的差异程度。结果表明,Group B基因序列保守碱基数比Group A、Group C、Group D的高,这是由于Group B上基因登录的植物分类纲级和基因类型一致,都是双子叶植物的GID1b型,因此比其他组的基因亲缘关系更近。Group A与Group B为旁系枝,Group A上分布的基因都是单子叶植物的GID1s,Group A基因数比Group B多,而基因序列的保守碱基数比Group B少,Group A基因间的亲缘关系比Group B远。Group C分布双子叶植物GID1a和GID1c基因,从图2可以看出,Group C基因主要分两大进化方向,上枝由27个GID1基因组成,下枝由31个基因组成,整体58个基因;上枝保守碱基数340,下枝基因的保守碱基数409,整体Group C的保守碱基数258。显示基因序列的保守碱基数与基因序列数成反比。Group D有10个基因,基因数比Group B多,但Group D基因的保守碱基数显著少于Group B(表2)。这个差异的原因是Group D分布2门植物的基因。Group A、Group B、Group C、Group D各组之间基因序列的保守碱基数差异显著,但98个基因有共同的保守序列。
3 討论
赤霉素受体GID1作为赤霉素信号传导途径的重要因子,最早在水稻[10]中获得,之后从拟南芥[11]、棉花[12]和大麦[13]等多种植物中获得,后续更多植物的GID1基因被克隆。从GID1基因的登录情况可以看出,被子植物门双子叶植物的GID1基因已被大量克隆登录;但单子叶植物、裸子植物和蕨类植物GID1基因的克隆甚少。单子叶植物的禾本科植物、薯蓣科植物应用广泛,主要应用于粮、饲料、药等。裸子植物是绿化树种中极为重要的一类[14],蕨类植物能监测大气重金属污染程度[15],苔藓结皮层能够改良土壤[16],这些植物在园林绿化中有着至关重要的地位。因此,克隆研究植物GID1基因可为研究其生长发育奠定基础,能充分发挥其重要的地位,至于更多单子叶植物、裸子植物和蕨类等植物的GID1基因有待克隆。
Yoshida等[5]和Gazara等[8]对GID1进行系统发育分析,发现几乎所有双子叶植物的基因都含有2种GID1,即GID1ac型和GID1b型。通过重建91个物种的GID1s系统发育树,发现GID1s在Group A单子叶植物中没有GID1a、GID1b、GID1c分类现象;在双子叶植物植物Group B和Group C中有简单的GID1b、GID1a和GID1c聚类现象,说明单子叶植物GID1基因型进化方向没有典型的基因分类现象,双子叶植物基因进化为典型的GID1b、GID1a和GID1c。与前者研究比较确定了双子叶植物存在GID1a,且GID1c和GID1b型进化方向,但单子叶植物的GID1基因的登录还不全,有待进一步研究。
在双子叶植物中,十字花科的拟南芥(Arabidopsis thaliana)、猕猴桃科的猕猴桃(Actinidia deliciosa)、 梧桐科的可可(Theobroma cacao)、茄科的番茄(Solanum lycopersicum)、 毛茛科的牡丹(Paeonia suffruticosa)、大戟科的橡胶(Hevea brasiliensis)和戟科的木薯(Manihot esculenta)已登录2个GID1基因。同样单子叶植物中,薯蓣科的山药(Dioscorea polystachya)、禾本科的玉米(Zea mays)、普通小麦(Triticum aestivum)和芭蕉科的小果野蕉(Musa acuminate)等植物也登录了2个GID1基因。猕猴桃、可可、玉米等2个基因为旁系枝自展值达90%以上,其他植物的基因非旁系枝,发现植物的基因之间不仅存在物种差异,还存在基因类型差异,比如猕猴桃的GID1a和GID1b之间基因同源性达到67%,山药的GID1a和GID1b之间基因同源性达到45%。双子叶植物和单子叶植物的GID1基因数量表明,植物中含有2个以上GID1基因,Gazara等[8]研究发现,每个取样组织中至少有1个基因在表达,除了以上11种植物的GID1基因已克隆并登录2个基因,其他植物的GID1基因有待进一步克隆分析。从98个GID1基因聚类分析可知,GID1基因的起源始尚不明确,在植物基因组中可能出现多次复制,导致基因组的分枝发散。91种植物的98个GID1基因具有相同的保守区域。Guan等[17]研究了GID1基因的同源性,发现可依赖基因之间的保守性来克隆未知基因。
4 结论
更多植物未知的GID1基因有待克隆。系统发育树中GID1拥有共同的根系,单子叶植物纲中没有分类现象,而双子叶植物纲中有分类现象,说明基因组中出现复制进化。所有已知的GID1基因序列具有相同的保守区域。
参考文献
[1] OLSZEWSKI N,SUN T P,GUBLER F.Gibberellin signaling:Biosynthesis,catabolism,and response pathways[J].Plant Cell,2002,14(S1):S61-S80.
[2] 李巧峡,张丽,王玉,等.赤霉素调控植物开花及花器官发育的研究进展[J].中国细胞生物学学报,2019,41(4):746-758.
[3] 陈敏.紫花苜蓿赤霉素受体基因MsGID1b的克隆及功能分析[D].北京:中国农业科学院,2017.
[4] 黄桃鹏,李媚娟,王睿,等.赤霉素生物合成及信号转导途径研究进展[J].植物生理学报,2015,51(8):1241-1247.
[5] YOSHIDA H,TANIMOTO E,HIRAI T,et al.Evolution and diversification of the plant gibberellin receptor GID1[J].PNAS,2018,115(33):E7844-E7853.
[6] GE W J,STEBER C M.Positive and negative regulation of seed germination by the Arabidopsis GA hormone receptors,GID1a,b,and c[J].Plant Direct,2018,2(9):1-11.
[7] UEGUCHI TANAKA M,ASHIKARI M,NAKAJIMA M,et al.Gibberellin Insensitive Dwarf 1 encodes a soluble receptor for gibberellin[J].Nature,2005,437(7059):693-698.
[8] GAZARA R K,MOHARANA K C,BELLIENY RABELO D,et al.Expansion and diversification of the gibberellin receptor GIBBERELLIN INSENSITIVE DWARF1(GID1)family in land plants[J].Plant Mol Biol,2018,97(4/5):435-449.
[9] THOMAS S G,RIEU I,STEBER C M.Gibberellin metabolism and signaling[J].Vitamins & Hormones,2005,72:289-338.
[10] AYA K,UEGUCHI TANAKA M,KONDO M,et al.Gibberellin modulates anther development in rice via the transcriptional regulation of GAMYB[J].The plant cell,2009,21(5):1453-1472.
[11] NAKAJIMA M,SHIMADA A,TAKASHI Y,et al.Identification and characterization of Arabidopsis gibberellin receptors[J].Plant J,2006,46(5):880-889.
[12] ALEMAN L,KITAMURA J,ABDEL MAGEED H,et al.Functional analysis of cotton orthologs of GA signal transduction factors GID1 and SLR1[J].Plant Mol Biol,2008,68(1/2):1-16.
[13] CHANDLER P M,HARDING C A,ASHTON A R,et al.Characterization of gibberellin receptor mutants of barley(Hordeum vulgare L.)[J].Mol Plant,2008,1(2):285-294.
[14] 張亚平.裸子植物在城市园林绿化中的应用[J].现代园艺,2017(16):127-128.
[15] 李琦,籍霞,王恩辉,等.苔藓植物对青岛市大气重金属污染的生物监测作用[J].植物学报,2014,49(5):569-577.
[16] 闫德仁,张胜男,黄海广,等.沙漠苔藓植物分解的土壤改良效应[J].水土保持研究,2020,27(3):225-229,237.
[17] GUAN Y R,XUE J Q,XUE Y Q,et al.Effect of exogenous GA3 on flowering quality,endogenous hormones,and hormone and flowering associated gene expression in forcing cultured tree peony(Paeonia suffruticosa)[J].J Integr Agric,2019,18(6):1295-1311.
猜你喜欢 赤霉素 素养导向的生物学论证式概念教学实践中学生物学(2020年9期)2020-12-28不同处理对藿香及雾灵香花芥种子萌发的影响南方农业·下旬(2019年9期)2019-01-093对普罗旺斯薰衣草种子萌发的影响">GA3对普罗旺斯薰衣草种子萌发的影响天津农业科学(2018年5期)2018-06-23葡萄花芽分化过程中遮阴对新梢内源激素的影响江苏农业科学(2017年13期)2017-09-28不同处理方法对黄芩种子萌发的影响安徽农业科学(2017年18期)2017-07-10赤霉素(GA3)对菠菜性别分化的影响及分子机制江苏农业科学(2017年5期)2017-04-15赤霉素的生物合成和信号转导研究进展教育教学论坛(2015年16期)2015-12-09不同时期喷施赤霉素对再生稻芽苗及产量的影响现代农业科技(2015年14期)2015-09-11超声波、赤霉素对南天竹种子萌发的影响江苏农业科学(2014年12期)2015-04-02大枣花期喷赤霉素可增产山西果树(2014年3期)2014-07-15